CICLO DE KREBS
El ciclo de Krebs (conocido también como ciclo de los ácidos tricarboxílicos o ciclo del ácido cítrico) es un ciclo metabólico de importancia fundamental en todas las células que utilizan oxígeno durante el proceso de respiración celular. En estos organismos aeróbicos, el ciclo de Krebs es el anillo de conjunción de las rutas metabólicas responsables de la degradación y desasimilación de los hidrocarburos, las grasas y las proteínas en anhídrido carbónico y agua, con la formación de energía química.
El ciclo de Krebs es
una ruta
metabólica anfibólica, ya que participa tanto en
procesos catabólicos como anabólicos. Este ciclo proporciona muchos precursores
para la producción de algunos aminoácidos,
como por ejemplo el cetoglutarato y el oxalacetato, así como otras moléculas
fundamentales para la célula.

LA TRANSFORMACIÓN DEL PIRUVATO EN ACETIL-COA MEDIANTE
DESCARBOXILACIÓN OXIDATIVA.

La descarboxilación
oxidativa del pirúvico es un paso anterior al propio ciclo de Krebs. Durante la
glicolisis en el citoplasma se produce el pirúvico, que pasa por una etapa de
transición para convertirse en acetil-CoA para que pueda entrar en el ciclo de
Krebs.
{kind=link}
ETAPAS DEL CICLO DE KREBS
REACCIÓN 1: CITRATO SINTASA (DE OXALACETATO A CITRATO)
La reacción es sumamente exoergónica (ΔG'°=-31.4 kJ/mol), motivo por el cual este paso es irreversible. El citrato producido por la enzima, además, es capaz de inhibir competitivamente la actividad de la enzima.
Incluso estando la reacción muy favorecida (porque es exoergónica), la citrato sintasa puede ser perfectamente regulada. Este aspecto tiene una notable importancia biológica, puesto que permite una completa regulación del ciclo de Krebs completo, convirtiendo a la enzima en una especie de marcapasos del ciclo.


REACCIÓN 2: ACONITASA (DE CITRATO A ISOCITRATO)

REACCIÓN 3: ISOCITRATO DESHIDROGENASA (DE ISOCITRATO A
OXOGLUTARATO)

REACCIÓN 4: Α-CETOGLUTARATO DESHIDROGENASA (DE
OXOGLUTARATO A SUCCINIL-COA)

REACCIÓN 5: SUCCINIL-COA SINTETASA (DE SUCCINIL-COA A
SUCCINATO)
La energía procedente del tioéster viene convertida en energía ligada a una unión fosfato. El primer paso de la reacción genera un nuevo intermediario a alta energía, conocido como succinil fosfato. Sucesivamente, una histidina presente en el sitio catalítico remueve el fosfato de la molécula glucídica, generando el producto succinato y una molécula de fosfohistidina, que dona velozmente el fosfato a un nucleósido difosfato, recargándolo a trifosfato. Se trata del único paso del ciclo de Krebs en el que se produce una fosforilación a nivel de sustrato.

REACCIÓN 6: SUCCINATO DESHIDROGENASA (DE SUCCINATO A
FUMARATO)

La primera reacción de oxidación es catalizada por el complejo enzimático de la succinato deshidrogenasa, la única enzima del ciclo que tiene como aceptor de hidrógeno al FAD en vez de al NAD+. El FAD es enlazado de modo covalente a la enzima por un residuo de histidina. La enzima se vale del FAD ya que la energía asociada a la reacción no es suficiente para reducir el NAD+.
REACCIÓN 7: FUMARASA (DE FUMARATO A L-MALATO)
REACCIÓN 8: MALATO DESHIDROGENASA (DE L-MALATO A OXALACETATO)
La energía libre de Gibbs asociada con esta última reacción es decididamente positiva, a diferencia de las otras del ciclo. La actividad de la enzima es remolcada por el consumo de oxalacetato por parte del citrato sintasa, y de NADH por parte de la cadena de transporte de electrones.
ENTRADAS Y SALIDAS.
En total, en el ciclo
de Krebs entran 1 molécula de acetil-CoA y 3 moléculas de H2O. Después de su
transcurso obtenemos:
1 molécula de
Coenzima A
3 NADH/H+ a
partir NAD+
1 molécula de GTP
(guanosina trofosfato) a partir de GDP + Pi
1 molécula de
Coenzima Q reducida (ubiquinol)
Los NADH/H+ y el
ubiquinol tienen un papel importante en la cadena respiratoria para la
producción de ATP, producto final de la respiración celular.
.

Las enzimas que
juegan un papel en el ciclo de Krebs y las reacciones que catalizan son las
siguientes:
1 -
La citrato sintetasa facilita la unión del oxalacetato
con el resto acílico que lleva la coenzima A. Para ello se necesita
adicionalmente un H2O y al final la coenzima A queda libre.
2 y 3 –
La aconitasa cataliza la producción de cis-aconitato quitándo un H2O
del citrato. Después incorpora un H2O al cis-aconitato para formar isocitrato.
4 –
La isocitrato deshidrogenasa oxida el isocitrato (y reduce al mismo
tiempo NAD+, produciendo NADH/H+). Como producto intermedio de este paso
resulta oxalosuccinato (no aparece en el esquema) que se convierte en
alfa-cetoglutarato mediante la descarboxilación. Resulta que el producto de
este paso contiene 5 átomos de carbono en vez de 6. El grupo carboxílico se
libera en forma de dióxido de carbono (CO2).
5 – El
alfa-cetoglutarato se une con una coenzima A con la ayuda de
la alfa-cetoglutarato-deshidrogenasa para formar succinil-CoA. En
este paso se libera otro CO2, lo que deja el producto con 4 átomos de carbono.
Además se genera un NADH/H+.
6 - Durante la
reacción 6 que es catalizada por la succinil-CoA-sintetasa, se genera el
succinato y una molécula de GTP (un compuesto rico en energía). La coenzima A
queda libre otra vez para reacciones siguientes.
7 –
La succinato-deshidrogenasa procede a la oxidación del succinato
formando el fumarato. En la misma reacción se obtiene un FADH2, que a
continuación reduce a la coenzima Q (ubiquinona), generando
QH2 (ubiquinol).
8 – Sigue la
hidratación del fumarato por la fumarasa y se obtiene el malato.
9 – Finalmente,
la malato-deshidrogenasa permite la oxidación del malato, generando
oxalacetato y otro NADH/H+. Regenerado, el oxalacetato puede aceptar de nuevo
un acetil-CoA y recorrer el ciclo, ganando más “energía” en forma de
NADH/H+ y QH2 que puede ser utilizada en la cadena respiratoria.
FUENTES CONSULTADAS:
·
https://es.wikipedia.org/wiki/Ciclo_de_Krebs
·
http://www.ciclodekrebs.com/
·
http://biologia.laguia2000.com/bioquimica/el-ciclo-de-krebs-paso-a-paso
·
http://es.slideshare.net/Regaladiux/ciclo-de-krebs-10063165
Realizado por: LORENA
HUERTAS GIL
CADENA RESPIRATORIA
Conceptos Generales
La misión de la
cadena transportadora de electrones es la de crear un gradiente electroquímico
que se utiliza para la síntesis de ATP. Dicho gradiente electroquímico se
consigue mediante el flujo de electrones entre diversas sustancias de esta
cadena que favorecen en último caso la translocación de protones que generan el
gradiente anteriormente mencionado. De esta forma podemos deducir la existencia
de tres procesos totalmente dependientes:
Un flujo de
electrones desde sustancias individuales
Un uso de la energía
desprendida de ese flujo de electrones que se utiliza para la translocación de
protones en contra de gradiente, por lo que energéticamente estamos hablando de
un proceso desfavorable.
Un uso de ese
gradiente electroquímico para la formación de ATP mediante un proceso favorable
desde un punto de vista energético.
En este punto la
célula ha ganado solo 4 ATP, 2 en la glucólisis y dos en el ciclo de Krebs, sin
embargo ha capturado electrones energéticos en 10 NADH2 y 2 FADH2. Estos
transportadores depositan sus electrones en el sistema de transporte de
electrones localizado en la membrana interna de la mitocondria.
{kind=link}
La cadena
respiratoria está formada por una serie de transportadores de electrones
situados en la cara interna de las crestas mitocondriales y que son capaces de
transferir los electrones procedentes de la oxidación del sustrato hasta el
oxígeno molecular, que se reducirá formándose agua.
Como resultado de
esta transferencia de electrones, los transportadores se oxidan y se reducen
alternativamente, liberándose una energía que en algunos casos es suficiente
para fosforilar el ADP y formar una molécula de ATP. Se trata de
la fosforilación oxidativa que permite ir almacenando en enlaces
ricos en energía la energía contenida en las moléculas NADH2, FADH2, NADPH2,
que se liberan en la glucólisis y en el ciclo de Krebs y que será más tarde
fácilmente utilizada. Toda cadena respiratoria que comience por el NAD conduce
a la formación de 3 ATP mientras que si comienza por el FAD produce sólo 2 ATP.
El rendimiento energético del NADP es similar al del NAD, así como el del GTP
lo es al del ATP.
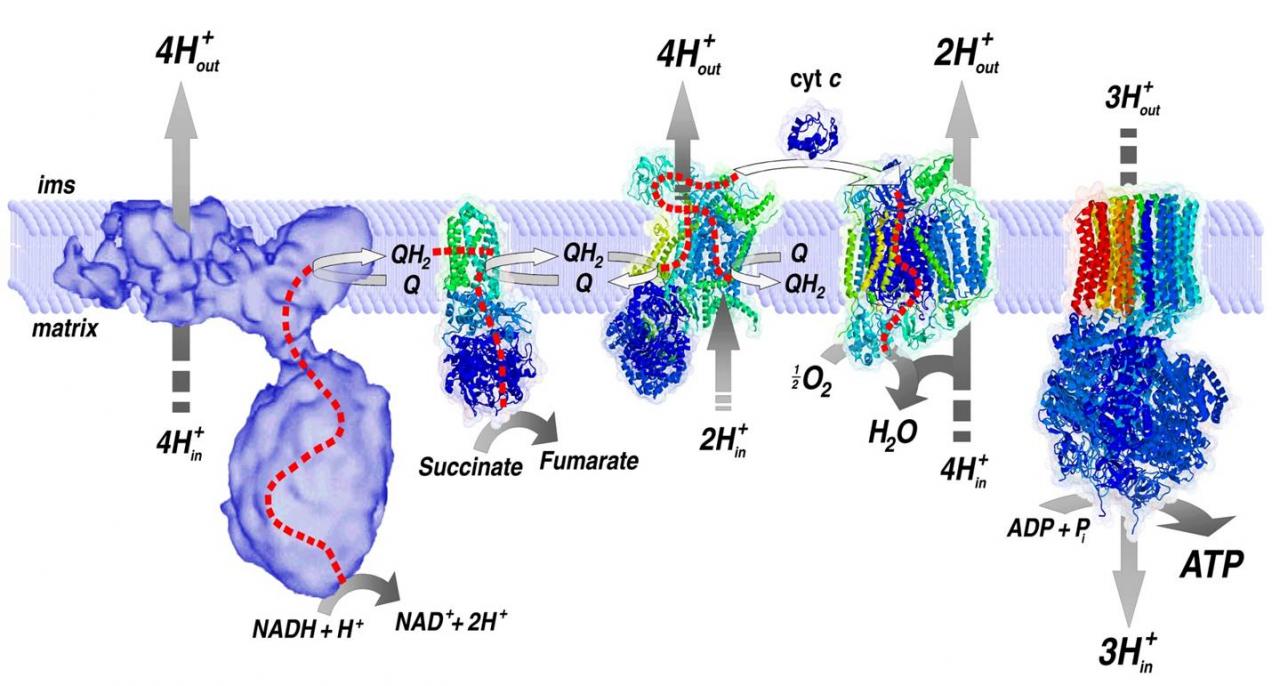
La cadena respiratoria mitocondrial consta de una serie de transportadores electrónicos que actúan secuencialmente, la mayoría de los cuales son proteínas integrales con grupos prostéticos capaces de aceptar y donar 1 o 2 electrones. Entre los principales componentes de la cadena de transporte de electrones se encuentran las moléculas conocidas como citocromos.
|
|
|
ATPasa bacteriana
|

Cada NADH2 que llega a la cadena respiratoria cede una pareja de electrones que en su transporte liberan energía suficiente para bombear 6 H+ desde la matriz al espacio intermembrana. Si los electrones proceden del FADH2 sólo se bombean 4 H+. Por cada 2 H+que vuelven a la matriz a través de la ATP-sintetasa se fosforila un ADP. Por tanto en la cadena respiratoria, se pueden obtener 3 ATP por cada NADH2 obtenido en el catabolismo y 2 ATP por cada FADH2.
CARACTERÍSTICAS DE LA CADENA RESPIRATORIA
La cadena
respiratoria mitocondrial o cadena de transporte de electrones está embebida en
la membrana interna mitocondrial, y la constituyen cinco complejos
multienzimáticos (I, II, III, IV y V o ATP sintasa) y dos transportadores de
electrones móviles (coenzima Q o ubiquinona y citocromo c).
Su principal función
es el trasporte coordinado de protones y electrones, para producir energía en
forma de ATP a partir de ADP y fosfato inorgánico. El transporte de electrones
genera energía que es utilizada para transportar protones de la matriz
mitocondrial al espacio intermembrana situado entre las membranas
mitocondriales externa e interna. Este proceso genera un gradiente
electroquímico de protones, que es utilizado por el complejo V (ATP sintasa)
para generar ATP a medida que los protones fluyen de nuevo desde el espacio
intermembrana a la matriz mitocondrial. El ATP generado es exportado al
citoplasma a través del transportador de nuecleótidos de adenina (ANT).

{kind=link}
EJEMPLO DE CADENA DE RESPIRACIÓN:
Glicólisis. En el
inicio del proceso de respiración. La glucosa traspasa la membrana celular y en
el citoplasma la molécula de glucosa sufre un proceso de oxidación,
dividiéndose en dos moléculas de ácido pirúvico, llamado también pirúvico.
Otras sustancias, como los aminoácidos, también se oxidan, desprendiéndose
aminas y pirúvico.
Descarboxilación. Las
moléculas de pirúvico entran a la mitocondria, donde comienzan a ser atacadas
por las enzimas que producen la descarboxilación oxidativa. Al principio una
enzima se encarga de desprender uno de los carbonos del ácido pirúvico
(desprendiendo CO2), y al mismo tiempo otra enzima se encarga de desprender dos
átomos de hidrógeno, produciéndose un radical Acetilo (ácido acético sin
radical hidroxilo -OH).
Ciclo de Krebs. Los
radicales acetilo son transportados por otra enzima, llamadas “Coenzima A”,
hacia la matriz mitocondrial (el núcleo de la mitocondria), donde los radicales
acetilo son oxidados y se libera energía. En esta fase también se recombinan
las moléculas de
CO2 con la coenzima
A para producir un total de 6 moléculas de acetilo oxidadas, que forman las
coenzimas NADH y FADH2.
En el siguiente paso,
las coenzimas resultantes, llamadas NADH y FADH2 son oxidadas nuevamente, para
que tengan electronegatividad y puedan aceptar electrones y protones, que son
proporcionados por otras coenzimas, que agregan electrones y fosforilan
(agregan fósforo) a las coenzimas, hasta agregar tres moléculas de fósforo y
moléculas de oxígeno, para producir el ATP.
FUENTES CONSULTADAS:
·
https://respiracioncelular.wordpress.com/cadena-respiratoria/
·
http://www.neurowikia.es/content/caracter%C3%ADsticas-de-la-cadena-respiratoria
·
http://es.slideshare.net/profesorjano/cadena-respiratoria
·
http://www.ejemplode.com/36-biologia/285-funciones_de_la_cadena_respiratoria.html
Realizado por: LORENA HUERTAS GIL
No hay comentarios.:
Publicar un comentario